Влияние полисахаридов из Atractylodes macrocephala и н-бутанола на митохондриальную аутофагию у крыс с дефицитом селезенки
Дефицит селезенки - это распространенный тип синдрома, связанный с нарушением функции органов и внутренних органов, обычно проявляющийся истощением, потерей аппетита, болями в эпигастральной области, вздутием живота, усталостью, бледным цветом лица, жидким стулом и т.д. Дефицит селезенки указывает на то, что организм находится в состоянии низкоэнергетического метаболизма, особенно связанного с дисфункцией желудочно-кишечного тракта. Стоит отметить, что митохондрии являются важным источником энергии для перистальтики желудочно-кишечного тракта в организме, производя энергию в виде АТФ посредством окислительного фосфорилирования, и являются важным звеном в "транспорте, опосредованном селезенкой". Окислительное повреждение, дефицит питательных веществ и другие внешние раздражители могут легко привести к повреждению митохондрий. В ответ на это клетки хозяина могут активировать механизмы аутофагии, чтобы очистить поврежденные митохондрии и поддержать клеточный гомеостаз. Если функция аутофагии нарушается и поврежденные митохондрии не могут быть своевременно очищены, это может привести к гибели клеток. Исследования показали, что нарушения митохондриальной аутофагии наблюдаются в таких клетках, как скелетные мышцы, нервы, печень и миокард у животных с дефицитом селезенки.
Atractylodes lancea (Thunb.) DC - многолетнее травянистое растение семейства Asteraceae, обладающее эффектом осушения сырости и укрепления селезенки. Согласно Китайской фармакопее, его источником является корневище растения Atractylodes macrocephala или Atractylodes macrocephala из семейства Asteraceae. Дикорастущие ресурсы Atractylodes macrocephala в основном распространены в Хубэй, Сюаньчэн, Аньхой, Вунин, Цзянси и Ханьчжун, Шэньси, а дикорастущие ресурсы Atractylodes macrocephala - в Северо-Восточном Китае, Внутренней Монголии, Хэбэе и других регионах. Традиционно считается, что Atractylodes macrocephala в районе Маошань провинции Цзянсу обладает наилучшим качеством и является настоящим лекарственным растением. Во многих странах, таких как Таиланд и Япония, Atractylodes macrocephala также широко используется для лечения заболеваний пищеварительной системы. Современные фармакологические исследования показали, что Atractylodes macrocephala может регулировать уровень желудочно-кишечных гормонов в сыворотке крови крыс; экстракт Atractylodes macrocephala оказывает ингибирующее действие на рост клеток рака желудка (BGC-823 и SGC-7901) и Helicobacter pylori. Atractylodes macrocephala содержит различные типы химических компонентов, включая летучие масла, полисахариды, гликозиды и низкополярные нелетучие вещества. Исследовательская группа ранее обнаружила, что полисахариды из Atractylodes macrocephala и его н-бутанольные компоненты (гликозиды) оказывают терапевтическое действие при дефиците селезенки. Исходя из этого, в данном эксперименте впервые изучается механизм действия эффективных компонентов Atractylodes macrocephala, регулирующих дефицит селезенки, с точки зрения митохондриальной аутофагии, с целью заложить основу для рационального применения Atractylodes macrocephala.




 Клинический диагноз недостаточности селезенки обычно основывается на симптомах. У пациентов с дефицитом селезенки часто наблюдаются такие симптомы, как снижение аппетита, тупые боли в животе, усталость и холодные конечности. У животных с дефицитом селезенки потеря аппетита проявляется в виде снижения веса, боль в животе - в виде горбатости, усталость и слабость - в виде вялости и легкой утомляемости, а боязнь холода - в виде скопления и скручивания. В этом эксперименте после моделирования у крыс наблюдались признаки истощения, горбатости, вялости и свернутости, что подтвердило успешность созданной модели крысы с дефицитом селезенки.
Клинический диагноз недостаточности селезенки обычно основывается на симптомах. У пациентов с дефицитом селезенки часто наблюдаются такие симптомы, как снижение аппетита, тупые боли в животе, усталость и холодные конечности. У животных с дефицитом селезенки потеря аппетита проявляется в виде снижения веса, боль в животе - в виде горбатости, усталость и слабость - в виде вялости и легкой утомляемости, а боязнь холода - в виде скопления и скручивания. В этом эксперименте после моделирования у крыс наблюдались признаки истощения, горбатости, вялости и свернутости, что подтвердило успешность созданной модели крысы с дефицитом селезенки.
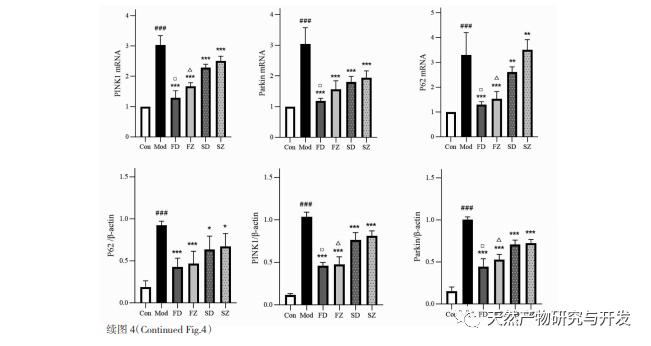
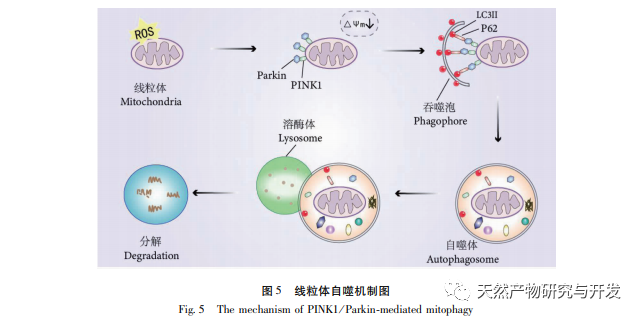
Механизм регуляции митохондриальной аутофагии сложен, и множество сигнальных путей могут опосредовать различные типы процессов аутофагии. Путь PINK1/Parkin является наиболее распространенным и хорошо изученным сигнальным путем аутофагии в митохондриях. PINK1 обычно не обнаруживается в здоровых митохондриях, поскольку после попадания в митохондрии он расщепляется внутримембранными гидролазами (Parl) и впоследствии очищается. Как показано на рисунке 5, при повреждении митохондрий вход PINK1 в митохондрии блокируется из-за снижения потенциала внутренней мембраны, он постоянно накапливается на внешней мембране митохондрий и рекрутирует паркин в поврежденные митохондрии. Паркин - это E3-убиквитин-протеин лигаза, которая может убиквитинировать поврежденные белки митохондриальной мембраны при активации PINK1. Он распознается селективным адаптерным белком аутофагии p62 и затем соединяется с белком, связанным с аутофагией, микротрубочковым ассоциированным белком 1 легкой цепи 3 (LC3) на мембране аутофагосомы, инициируя аутофагию и в конечном итоге разрушая поврежденные митохондрии. В ходе исследования было установлено, что уровни мРНК и белка PINK1 и паркина значительно увеличились в группе модели, что свидетельствует о наличии поврежденных митохондрий. Путь PINK1/Parkin был активирован, но поврежденные митохондрии, возможно, не были деградированы аутофагией из-за дисфункции. После обработки полисахаридами Atractylodes macrocephala и н-бутанолом уровни мРНК и белков PINK1 и Parkin в каждой группе крыс снизились, что указывает на то, что некоторые поврежденные митохондрии были деградированы и очищены аутофагией.
Митохондриальная аутофагия тесно регулируется многими белками, связанными с аутофагией, поэтому для определения силы аутофагии часто используют экспрессию некоторых ключевых белков, среди которых LC3 и p62 являются более важными. Когда аутофагия не происходит, большая часть LC3 существует в цитоплазме в форме LC3I. Когда аутофагия происходит, LC3 подвергается модификации убиквитинирования и связывается с фосфатидилэтаноламином на поверхности аутофагосом, образуя LC3II, расположенный на поверхности мембраны аутофагосом. Поэтому появление LC3II часто является важным маркером аутофагии. Как ключевой адаптерный белок селективной аутофагии, p62 может связываться с LC3II и служить субстратом для селективной аутофагии. Когда лизосомы и аутофагосомы находятся на стадии связывания, p62 может выступать в качестве донора для лизосом, направляя аутофагосомы на связывание с ними и формирование аутолизосом. Наконец, субстраты деградируют под действием протеолитических ферментов в лизосомах, и p62 впоследствии деградирует вместе с ними. Аутофагия обычно происходит при снижении уровня p62. В данном исследовании было отмечено, что экспрессия белка LC3II и соотношение LC3II/I в группе моделирования были значительно ниже, чем в группе сравнения, что указывает на ограничение образования аутофагосом в клетках антральной ткани желудка при дефиците селезенки. Повышение уровня мРНК и белка p62 указывает на ингибирование активности аутофагии. После введения полисахаридов Atractylodes macrocephala, Atractylodes macrocephala и н-бутанола в каждой группе крыс усилилось преобразование LC3 Ⅰ в LC3II, а уровни мРНК и белка p62 снизились, что указывает на повышение уровня аутофагии. Согласно результатам просвечивающей электронной микроскопии, структура митохондрий у крыс модельной группы была сильно повреждена, а аутофагической структуры не наблюдалось, что указывает на то, что нарушение функции аутофагии могло привести к накоплению поврежденных митохондрий. После введения препарата структурные повреждения митохондрий в каждой группе крыс уменьшились, а количество аутофагических лизосом увеличилось. Таким образом, мы считаем, что дефицит селезенки приводит к нарушению функции митохондриальной аутофагии в антральной ткани желудка крыс, а введение полисахаридов из Atractylodes macrocephala и Atractylodes macrocephala с н-бутанолом способствует развитию аутофагии.
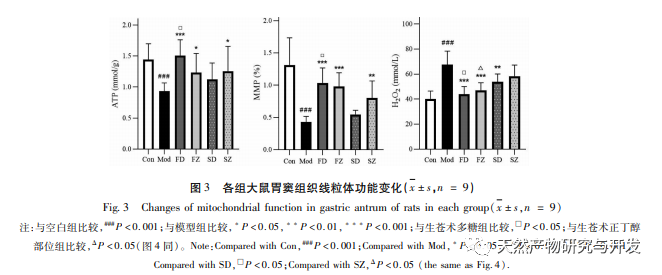
Нарушение митохондриальной аутофагии обычно сопровождается выработкой реактивных форм кислорода (ROS), а повышение уровня внутриклеточных ROS провоцирует окислительный стресс. Окислительный стресс поражает мембрану митохондрий, что приводит к снижению мембранного потенциала, повреждению митохондрий и функциональным нарушениям, которые, в свою очередь, вызывают дефекты выработки энергии и повышение уровня ROS, создавая порочный круг. Поврежденные и дисфункциональные митохондрии избирательно очищаются митохондриальной аутофагией, которая помогает снизить выработку ROS и предотвратить окислительное повреждение нормальных клеток. H2O2 является основным эндогенным ROS в клетках, который может вырабатываться почти всеми окислительными стрессорами и свободно диффундировать в клетки и из них, вызывая дисбаланс в окислительной антиоксидантной системе. Поэтому в данном эксперименте уровень РОС оценивается косвенно, путем определения содержания H2O2. Результаты исследования показали, что уровень H2O2, мембранного потенциала и АТФ у крыс с дефицитом селезенки снизился, что свидетельствует о нарушении функции митохондрий. Лечение полисахаридами из Atractylodes macrocephala и н-бутанолом значительно улучшило функцию митохондрий, что может быть достигнуто за счет стимулирования митохондриальной аутофагии и увеличения клиренса поврежденных митохондрий.
Таким образом, мы предполагаем, что полисахариды и н-бутаноловые компоненты Atractylodes macrocephala могут регулировать дефицит селезенки путем улучшения аутофагии в тканях антрума желудка и восстановления функции митохондрий. Более того, полисахариды и н-бутанольные компоненты после обжаривания с отрубями значительно эффективнее, чем сырой продукт.