Efectos de los polisacáridos de Atractylodes macrocephala y del n-butanol sobre la autofagia mitocondrial en ratas con deficiencia de bazo
La deficiencia de bazo es un tipo de síndrome común en la diferenciación de órganos y vísceras, que suele manifestarse como emaciación, pérdida de apetito, dolor epigástrico, hinchazón, fatiga, tez pálida, heces blandas, etc. La deficiencia de bazo indica que el organismo se encuentra en un estado metabólico de baja energía, especialmente relacionado con la disfunción gastrointestinal. Cabe señalar que las mitocondrias son una importante fuente de energía para el peristaltismo gastrointestinal en el organismo, produciendo energía en forma de ATP mediante la fosforilación oxidativa, y son un eslabón importante en el "transporte mediado por el bazo". El daño oxidativo, la deficiencia de nutrientes y otros estímulos externos pueden provocar fácilmente daños mitocondriales. En respuesta, las células huésped pueden activar mecanismos de autofagia para eliminar las mitocondrias dañadas y mantener la homeostasis celular. Si la función de la autofagia se deteriora y las mitocondrias dañadas no pueden eliminarse a tiempo, puede inducirse la muerte celular. La investigación ha descubierto que se han observado anomalías de la autofagia mitocondrial en células como el músculo esquelético, los nervios, el hígado y el miocardio en animales con deficiencia de bazo.
Atractylodes lancea (Thunb.) DC es una planta herbácea perenne de la familia Asteraceae, que tiene el efecto de secar la humedad y fortalecer el bazo. La Farmacopea China estipula que su fuente es el rizoma de la planta Asteraceae Atractylodes macrocephala o Atractylodes macrocephala. Los recursos silvestres de Atractylodes macrocephala se distribuyen principalmente en Hubei, Xuancheng, Anhui, Wuning, Jiangxi, y Hanzhong, Shaanxi, mientras que los recursos silvestres de Atractylodes macrocephala se distribuyen principalmente en el noreste de China, Mongolia Interior, Hebei y otras regiones. Tradicionalmente, se cree que la Atractylodes macrocephala de la zona de Maoshan, en la provincia de Jiangsu, tiene la mejor calidad y es una hierba medicinal genuina. En muchos países, como Tailandia y Japón, la Atractylodes macrocephala también se utiliza habitualmente para tratar enfermedades del aparato digestivo. Estudios farmacológicos modernos han demostrado que la Atractylodes macrocephala puede regular los niveles de hormonas gastrointestinales en el suero de ratas; el extracto de Atractylodes macrocephala tiene efectos inhibidores sobre el crecimiento de células de cáncer gástrico (BGC-823 y SGC-7901) y Helicobacter pylori. Atractylodes macrocephala contiene varios tipos de componentes químicos, como aceites volátiles, polisacáridos, glucósidos y sustancias no volátiles de baja polaridad. El equipo de investigación descubrió anteriormente que los polisacáridos de Atractylodes macrocephala y sus componentes n-butanol (glucósidos) tienen efectos terapéuticos sobre la deficiencia de bazo. Basándose en esto, este experimento es el primero en explorar el mecanismo de los componentes eficaces de Atractylodes macrocephala que regulan la deficiencia de bazo desde la perspectiva de la autofagia mitocondrial, con el objetivo de sentar las bases para la aplicación racional de Atractylodes macrocephala.




 El diagnóstico clínico de la deficiencia de bazo suele basarse en los síntomas. Los pacientes con deficiencia de bazo suelen presentar síntomas como disminución del apetito, dolor abdominal sordo, fatiga y extremidades frías. La pérdida de apetito se manifiesta como pérdida de peso en los animales con deficiencia de bazo, el dolor abdominal se manifiesta como joroba, la fatiga y la debilidad se manifiestan como apatía y fatiga fácil, y el miedo al frío se manifiesta como agrupamiento y acurrucamiento. En este experimento, las ratas mostraron signos de emaciación, joroba, apatía y acurrucamiento tras el modelado, lo que confirmó el éxito del modelo de rata con deficiencia de bazo establecido.
El diagnóstico clínico de la deficiencia de bazo suele basarse en los síntomas. Los pacientes con deficiencia de bazo suelen presentar síntomas como disminución del apetito, dolor abdominal sordo, fatiga y extremidades frías. La pérdida de apetito se manifiesta como pérdida de peso en los animales con deficiencia de bazo, el dolor abdominal se manifiesta como joroba, la fatiga y la debilidad se manifiestan como apatía y fatiga fácil, y el miedo al frío se manifiesta como agrupamiento y acurrucamiento. En este experimento, las ratas mostraron signos de emaciación, joroba, apatía y acurrucamiento tras el modelado, lo que confirmó el éxito del modelo de rata con deficiencia de bazo establecido.
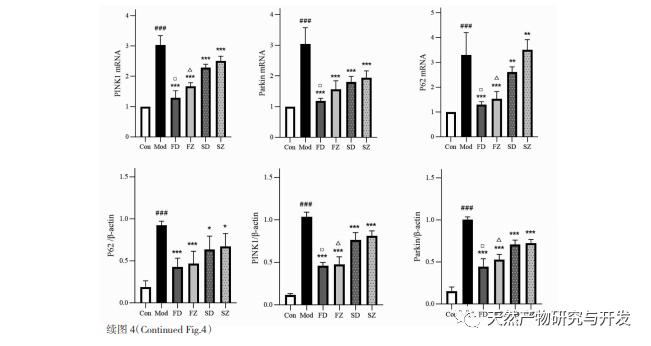
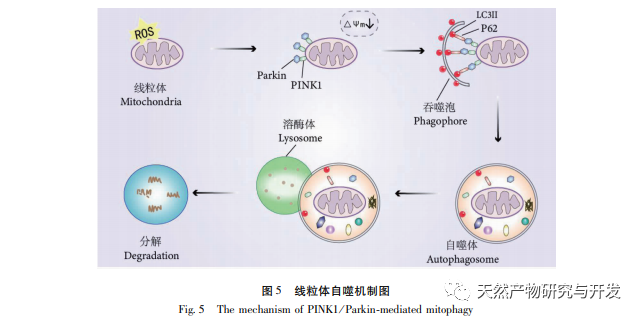
El mecanismo regulador de la autofagia mitocondrial es complejo, y múltiples vías de señalización pueden mediar diferentes tipos de procesos de autofagia. La vía PINK1/Parkin es la vía de señalización de la autofagia más común y mejor estudiada en las mitocondrias. PINK1 suele ser indetectable en mitocondrias sanas porque es escindida por hidrolasas intramembrana (Parl) tras entrar en la mitocondria y posteriormente eliminada. Como se muestra en la Figura 5, cuando las mitocondrias están dañadas, la entrada de PINK1 en la vía mitocondrial se bloquea debido a una disminución del potencial de la membrana interna, acumulándose continuamente en la membrana externa de las mitocondrias y reclutando parkina a las mitocondrias dañadas. Parkin es una proteína ligasa ubiquitina E3 que puede ubiquitinar proteínas de membrana mitocondrial dañadas cuando es activada por PINK1. Es reconocida por la proteína adaptadora selectiva de la autofagia p62 y luego se conecta a la proteína relacionada con la autofagia proteína asociada a microtúbulos 1 cadena ligera 3 (LC3) en la membrana del autofagosoma, iniciando la autofagia y, en última instancia, degradando las mitocondrias dañadas. Este estudio descubrió que los niveles de ARNm y proteína de PINK1 y Parkin aumentaban significativamente en el grupo modelo, lo que indicaba la presencia de mitocondrias dañadas. La vía PINK1/Parkin se ha activado, pero es posible que las mitocondrias dañadas no hayan sido degradadas por la autofagia debido a su disfunción. Tras el tratamiento con polisacáridos de Atractylodes macrocephala y n-butanol, los niveles de ARNm y proteína de PINK1 y Parkin en cada grupo de ratas disminuyeron, lo que indica que algunas mitocondrias dañadas han sido degradadas y limpiadas por la autofagia.
La autofagia mitocondrial está estrechamente regulada por muchas proteínas relacionadas con la autofagia, por lo que la expresión de ciertas proteínas clave se utiliza a menudo para determinar la fuerza de la autofagia, entre las que LC3 y p62 son más críticas. Cuando no se produce autofagia, la mayor parte de LC3 existe en el citoplasma en forma de LC3I. Cuando se produce la autofagia, LC3 sufre una modificación por ubiquitinación y se une a la fosfatidiletanolamina en la superficie de los autofagosomas para formar LC3II localizada en la superficie de la membrana de los autofagosomas. Por lo tanto, la aparición de LC3II suele ser un marcador importante de autofagia. Como proteína adaptadora clave de la autofagia selectiva, p62 puede unirse a LC3II y servir como sustrato para la autofagia selectiva. Cuando los lisosomas y los autofagosomas están en fase de unión, p62 puede actuar como donante para los lisosomas, guiando a los autofagosomas para que se unan a ellos y formen autolisosomas. Finalmente, los sustratos son degradados por enzimas proteolíticas en los lisosomas, y p62 se degrada posteriormente junto con ellos. La autofagia suele producirse con una disminución de p62. En este estudio se observó que la expresión de la proteína LC3II y la relación LC3II/I en el grupo modelo eran significativamente inferiores a las del grupo blanco, lo que indica que la formación de autofagosomas en las células del tejido antral gástrico está restringida durante la deficiencia de bazo. El aumento de los niveles de ARNm y proteína p62 indica la inhibición de la actividad de autofagia. Tras la administración de polisacáridos de Atractylodes macrocephala, Atractylodes macrocephala y n-butanol, aumentó la conversión de LC3 Ⅰ a LC3II en cada grupo de ratas, y disminuyeron los niveles de ARNm y proteína p62, lo que indica un aumento de los niveles de autofagia. En correspondencia con los resultados de la microscopía electrónica de transmisión, la estructura mitocondrial de las ratas del grupo modelo estaba gravemente dañada, y no se observó ninguna estructura autofágica, lo que indica que el deterioro de la función de autofagia puede haber conducido a la acumulación de mitocondrias dañadas. Tras la administración, se redujo el daño estructural mitocondrial en cada grupo de ratas, y aumentó el número de lisosomas autofágicos. Por lo tanto, creemos que la deficiencia de bazo conduce a un deterioro de la función de autofagia mitocondrial en el tejido antral gástrico de rata, mientras que la administración de polisacáridos de Atractylodes macrocephala y Atractylodes macrocephala con n-butanol promueve la autofagia.
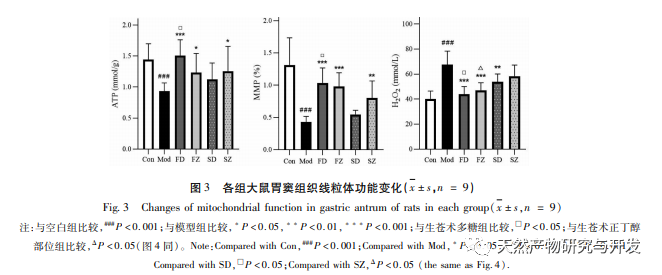
La obstrucción de la autofagia mitocondrial suele ir acompañada de la producción de especies reactivas del oxígeno (ROS), y un aumento de los niveles intracelulares de ROS inducirá estrés oxidativo. El estrés oxidativo ataca la membrana mitocondrial, provocando una disminución del potencial de membrana, daño mitocondrial y deterioro funcional, lo que a su vez desencadena defectos en la generación de energía y un aumento de ROS, creando un círculo vicioso. Las mitocondrias dañadas y disfuncionales son eliminadas selectivamente por la autofagia mitocondrial, que ayuda a reducir la producción de ROS y a prevenir el daño oxidativo a las células normales. El H2O2 es la principal ERO endógena en las células, que puede ser producida por casi todos los factores de estrés oxidativo y puede difundirse libremente dentro y fuera de las células, causando un desequilibrio en el sistema antioxidante oxidativo. Por lo tanto, este experimento evalúa indirectamente los niveles de ROS mediante la detección del contenido de H2O2. Los resultados de la investigación mostraron que los niveles de H2O2, potencial de membrana y ATP en ratas con deficiencia de bazo disminuyeron, lo que indica un deterioro de la función mitocondrial. El tratamiento con polisacáridos de Atractylodes macrocephala y n-butanol mejoró significativamente la función mitocondrial, lo que puede lograrse promoviendo la autofagia mitocondrial y aumentando el aclaramiento de las mitocondrias dañadas.
En resumen, especulamos que los polisacáridos y los componentes de n-butanol de Atractylodes macrocephala pueden regular la deficiencia de bazo mejorando los trastornos de autofagia en el tejido del antro gástrico y restaurando la función mitocondrial. Además, los polisacáridos y los componentes de n-butanol tras la fritura con salvado son significativamente más eficaces que el producto crudo.