The effects of polysaccharides from Atractylodes macrocephala and n-butanol on mitochondrial autophagy in spleen deficient rats
Spleen deficiency is a common syndrome type in the differentiation of organs and viscera, usually manifested as emaciation, loss of appetite, epigastric pain, bloating, fatigue, pale complexion, loose stools, etc. Spleen deficiency indicates that the body is in a low-energy metabolic state, especially related to gastrointestinal dysfunction. It is worth noting that mitochondria are an important energy source for gastrointestinal peristalsis in the body, producing energy in the form of ATP through oxidative phosphorylation, and are an important link in the “spleen mediated transport”. Oxidative damage, nutrient deficiency, and other external stimuli can easily lead to mitochondrial damage. In response, host cells can activate autophagy mechanisms to clear damaged mitochondria and maintain cellular homeostasis. If autophagy function is impaired and damaged mitochondria cannot be cleared in a timely manner, it can induce cell death. Research has found that mitochondrial autophagy abnormalities have been observed in cells such as skeletal muscle, nerves, liver, and myocardium in animals with spleen deficiency.
Atractylodes lancea (Thunb.) DC is a perennial herbaceous plant in the Asteraceae family, which has the effect of drying dampness and strengthening the spleen. The Chinese Pharmacopoeia stipulates that its source is the rhizome of the Asteraceae plant Atractylodes macrocephala or Atractylodes macrocephala. The wild resources of Atractylodes macrocephala are mainly distributed in Hubei, Xuancheng, Anhui, Wuning, Jiangxi, and Hanzhong, Shaanxi, while the wild resources of Atractylodes macrocephala are mainly distributed in Northeast China, Inner Mongolia, Hebei, and other regions. Traditionally, it is believed that Atractylodes macrocephala in the Maoshan area of Jiangsu Province has the best quality and is a genuine medicinal herb. In many countries such as Thailand and Japan, Atractylodes macrocephala is also commonly used to treat digestive system diseases. Modern pharmacological studies have shown that Atractylodes macrocephala can regulate the levels of gastrointestinal hormones in rat serum; Atractylodes macrocephala extract has inhibitory effects on the growth of gastric cancer cells (BGC-823 and SGC-7901) and Helicobacter pylori. Atractylodes macrocephala contains various types of chemical components, including volatile oils, polysaccharides, glycosides, and low polarity non-volatile substances. The research team previously found that polysaccharides from Atractylodes macrocephala and its n-butanol components (glycosides) have therapeutic effects on spleen deficiency. Based on this, this experiment is the first to explore the mechanism of Atractylodes macrocephala effective components regulating spleen deficiency from the perspective of mitochondrial autophagy, aiming to lay the foundation for the rational application of Atractylodes macrocephala.




 The clinical diagnosis of spleen deficiency is generally based on symptoms. Patients with spleen deficiency often exhibit symptoms such as decreased appetite, dull abdominal pain, fatigue, and cold limbs. Appetite loss is manifested as weight loss in animals with spleen deficiency, abdominal pain is manifested as hunchback, fatigue and weakness are manifested as listlessness and easy fatigue, and fear of cold is manifested as clustering and curling up. In this experiment, the rats showed signs of emaciation, hunchback, listlessness, and curled up after modeling, which confirmed the success of the spleen deficiency rat model established.
The clinical diagnosis of spleen deficiency is generally based on symptoms. Patients with spleen deficiency often exhibit symptoms such as decreased appetite, dull abdominal pain, fatigue, and cold limbs. Appetite loss is manifested as weight loss in animals with spleen deficiency, abdominal pain is manifested as hunchback, fatigue and weakness are manifested as listlessness and easy fatigue, and fear of cold is manifested as clustering and curling up. In this experiment, the rats showed signs of emaciation, hunchback, listlessness, and curled up after modeling, which confirmed the success of the spleen deficiency rat model established.
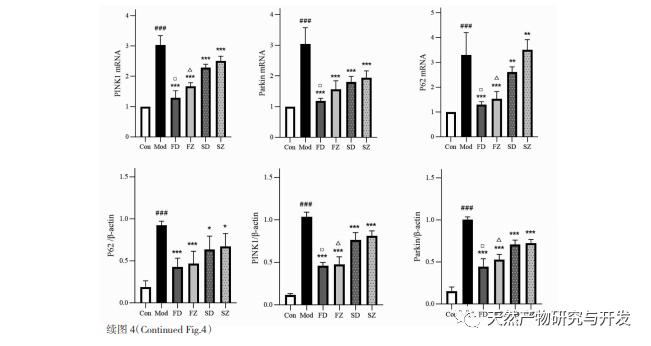
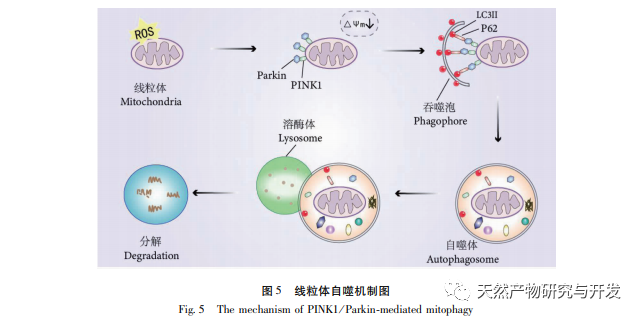
The regulatory mechanism of mitochondrial autophagy is complex, and multiple signaling pathways can mediate different types of autophagy processes. The PINK1/Parkin pathway is the most common and well studied autophagy signaling pathway in mitochondria. PINK1 is usually undetectable in healthy mitochondria because it is cleaved by intramembrane hydrolases (Parl) after entering the mitochondria and subsequently cleared. As shown in Figure 5, when mitochondria are damaged, PINK1’s entry into the mitochondrial pathway is blocked due to a decrease in inner membrane potential, continuously accumulating on the outer membrane of mitochondria and recruiting parkin to the damaged mitochondria. Parkin is an E3 ubiquitin protein ligase that can ubiquitinate damaged mitochondrial membrane proteins when activated by PINK1. It is recognized by the selective autophagy adaptor protein p62 and then connects to the autophagy related protein microtubule associated protein 1 light chain 3 (LC3) on the autophagosome membrane, initiating autophagy and ultimately degrading damaged mitochondria. This study found that the mRNA and protein levels of PINK1 and Parkin significantly increased in the model group, indicating the presence of damaged mitochondria. The PINK1/Parkin pathway has been activated, but damaged mitochondria may not have been degraded by autophagy due to dysfunction. After treatment with polysaccharides from Atractylodes macrocephala and n-butanol, the mRNA and protein levels of PINK1 and Parkin in each group of rats decreased, indicating that some damaged mitochondria have been degraded and cleared by autophagy.
Mitochondrial autophagy is closely regulated by many autophagy related proteins, so the expression of certain key proteins is often used to determine the strength of autophagy, among which LC3 and p62 are more critical. When autophagy does not occur, most LC3 exists in the cytoplasm in the form of LC3I. When autophagy occurs, LC3 undergoes ubiquitination modification and binds to phosphatidylethanolamine on the surface of autophagosomes to form LC3II located on the membrane surface of autophagosomes. Therefore, the appearance of LC3II is often an important marker of autophagy. As a key selective autophagy adapter protein, p62 can bind to LC3II and serve as a substrate for selective autophagy. When lysosomes and autophagosomes are in the binding stage, p62 can act as a donor for lysosomes, guiding autophagosomes to bind with them and form autolysosomes. Finally, substrates are degraded by proteolytic enzymes in lysosomes, and p62 is subsequently degraded together. Autophagy usually occurs with a decrease in p62. This study observed that the expression of LC3II protein and the LC3II/I ratio in the model group were significantly lower than those in the blank group, indicating that autophagosome formation in gastric antral tissue cells is restricted during spleen deficiency. The increase in p62 mRNA and protein levels indicates inhibition of autophagy activity. After administration of polysaccharides from Atractylodes macrocephala, Atractylodes macrocephala, and n-butanol, the conversion of LC3 Ⅰ to LC3II in each group of rats was enhanced, and the levels of p62 mRNA and protein decreased, indicating an increase in autophagy levels. Corresponding to the transmission electron microscopy results, the mitochondrial structure of the model group rats was severely damaged, and no autophagic structure was observed, indicating that impaired autophagy function may have led to the accumulation of damaged mitochondria. After administration, the mitochondrial structural damage in each group of rats was reduced, and the number of autophagic lysosomes increased. Therefore, we believe that spleen deficiency leads to impaired mitochondrial autophagy function in rat gastric antral tissue, while administration of polysaccharides from Atractylodes macrocephala and Atractylodes macrocephala with n-butanol promotes autophagy.
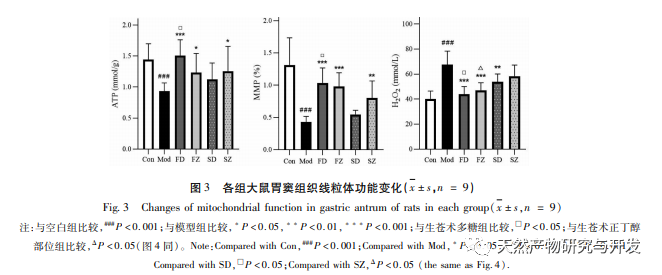
Mitochondrial autophagy obstruction is usually accompanied by the production of reactive oxygen species (ROS), and an increase in intracellular ROS levels will induce oxidative stress. Oxidative stress attacks the mitochondrial membrane, leading to a decrease in membrane potential, mitochondrial damage, and functional impairment, which in turn triggers energy generation defects and an increase in ROS, creating a vicious cycle. Damaged and dysfunctional mitochondria are selectively cleared by mitochondrial autophagy, which helps reduce ROS production and prevent oxidative damage to normal cells. H2O2 is the main endogenous ROS in cells, which can be produced by almost all oxidative stressors and can freely diffuse in and out of cells, causing an imbalance in the oxidative antioxidant system. Therefore, this experiment indirectly evaluates ROS levels by detecting H2O2 content. The research results showed that the levels of H2O2, membrane potential, and ATP in spleen deficient rats decreased, indicating impaired mitochondrial function. The treatment with polysaccharides from Atractylodes macrocephala and n-butanol significantly improved mitochondrial function, which may be achieved by promoting mitochondrial autophagy and increasing the clearance of damaged mitochondria.
In summary, we speculate that the polysaccharides and n-butanol components of Atractylodes macrocephala may regulate spleen deficiency by improving autophagy disorders in gastric antrum tissue and restoring mitochondrial function. Moreover, the polysaccharides and n-butanol components after stir frying with bran are significantly more effective than the raw product.